On Yeast And ALS (Part 1)
- 3 minsAlthough yeast and humans are separated by billions of years of evolution, yeast makes a really good disease model and can be really helpful to screen compounds and see which of them can rescue the phenotype acquired by inducing a mutation associated with our disease of interest. Below I expand on the 3 yeast models of ALS (actual yeast work to come during the summer!)
SOD1
SOD1 is a gene responsible for about 20% of fALS cases. Over 150 mutations passed through mendelian inheritance have been found in ALS (most autosomal dominant). Aggregates containing SOD1 are found in patients with familial ALS, but not in patients with sporadic ALS
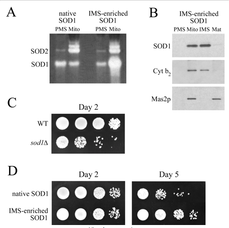
A model of SOD1 pathology in yeast was published by Sturtz, LA et al (2001)1. They found that knocking out SOD1 caused metabolic defects and rapid death.
TDP43
TDP43 is responsible for about 5% of fALS cases. Unlike SOD1, aggregates containing TDP43 are also found in sporadic ALS.
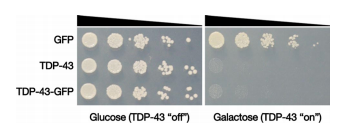
In a paper by Johnson, BS et al (2008) a TDP43 yeast model is described. TDP43 fused with GFP put under the control of a galactose promoter was induced in the yeast cells. Expression of TDP43 caused intranuclear aggregates, consistent with the TDP43 ALS phenotype. Increasing TDP43 expression caused the aggregates to move to the cytoplasm, Expressing TDP43 using galactose caused growth inhibition.
FUS
FUS like TDP43 is responsible for about 5% of fALS cases and it is found in both sporadic and familial ALS. FUS mutations in fALS are usually clustered around the extreme C terminus. Papers by Sun, Z et al (2010) and Ju, S et al (2011) describe yeast models of FUS toxicity.
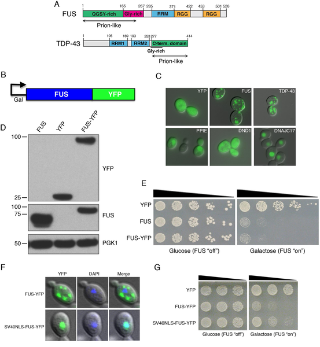
In the Ju paper they generated 3 strands of yeast: one with wild type FUS and the other two with mutant FUS (R521G and H517Q) fused to GFP and with the GAL1 promoter. FUS was found to be toxic to yeast (caused growth inhibition) in a dose-dependent manner. When either mutant or WT FUS is over expressed (with 2% galactose) aggregates in the cytoplasm are formed, consistent with the ALS phenotype.
Footnotes
1) I am not completely satisfied with this model, as it is thought ALS-linked mutations in SOD1 are likely to cause a toxic gain of function, not a loss of function. Given that, it would probably be better to make a yeast model containing one of the associated mutations rather than knocking out SOD1. I do not know if this has been done in the past, but if it has I was unable to find it.
References
Sturtz, LA et al. A Fraction of Yeast Cu,Zn-Superoxide Dismutase and Its Metallochaperone, CCS, Localize to the Intermembrane Space of Mitochondria: A PHYSIOLOGICAL ROLE FOR SOD1 IN GUARDING AGAINST MITOCHONDRIAL OXIDATIVE DAMAGE. J. Biol. Chem. 276, 38084-38089 (2001)
Bastow, EL et al. Using yeast models to probe the molecular basis of amyotrophic lateral sclerosis. Biochem Soc Trans. 39(5) 1482-1487 (2011)
Johnson, BS et al. A yeast TDP-43 proteinopathy model: Exploring
the molecular determinants of TDP-43 aggregation and cellular toxicity. PNAS, 105(17) 6439–6444 (2008)
Sun, Z et al. Molecular Determinants and Genetic Modifiers of Aggregation and Toxicity for the ALS Disease Protein FUS/TLS. PLoS Biology, E1000614-E1000614 (2010)
Ju, S et al. A Yeast Model of FUS/TLS-Dependent Cytotoxicity. PLoS Biology, E1001052-E1001052 (2011)

Jamie Cayley
Grad Student & TA at Missouri State University